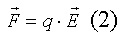Главная >> Биофизика Лекция №6. Электрический потенциал в биологических системах Электрический зарядЭлектрический заряд, подобно массе, является фундаментальным свойством веществ. Существует два типа зарядов, условно обозначенные как положительный и отрицательный.
Каждое вещество имеет электрический заряд, величина которого может быть положительной, отрицательной или быть равной нулю. Например, электроны заряжены отрицательно, а протоны - положительно. Поскольку каждый атом содержит один или более электронов и равное количество протонов, общее число зарядов в макроскопическом объекте - чрезвычайно большое, но в целом такой объект не заряжен или имеет небольшой заряд.
Заряд электрона является по абсолютной величине самым маленьким.
Электрическое поле. Закон КулонаКаждый заряженный объект образует в окружающем его пространстве электрическое поле. Электрическое поле является видом материи, посредством которой заряженные объекты взаимодействуют друг с другом. Пробный заряд, внесённый в электрическое поле другого заряда "чувствует" присутствие этого поля. Он будет притягиваться к заряду, создающему электрическое поле, или отталкиваться от него.
Закон Кулона определяет электрическую силу F, действующую между двумя точечными зарядами q1 и q2:
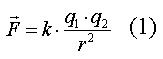 |
k - константа, определяемая выбранными условиями; r - расстояние между зарядами.
Согласно закону Кулона, сила действует в направлении линии, соединяющей два заряда. Величина силы, действующей на заряды, пропорциональна величине каждого из зарядов и обратно пропорциональна квадрату расстояния между ними.
Электрическое поле можно представить в виде силовых линий, показывающих направление электрических сил. Эти силы направлены от заряда, когда он положительный, и к заряду, если он отрицательный. Если положительный заряд поместить в электрическое поле, он подвергается действию силы в направлении поля. Отрицательный заряд подвергается силе, направленной противоположно направлению поля.
Характеристики электрического поля1) Напряжённость электрического поля. Каждый электрический заряд производит вокруг себя электрическое поле. Если другой заряд q внести в это поле, то на него будет действовать сила F, пропорциональная q и напряжённости электрического поля E:
Напряжённость электрического поля E (или просто напряжённость) в любой точке определяется как электрическая сила F, которая действует на положительной заряд q, помещённый в эту точку:
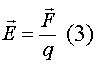 |
E - векторная величина, то есть имеет как величину, так и направление. Единицей измерения напряжённости является вольт на метр [В/м].
Принцип наложения (суперпозиции) указывает, что если электрическое поле создают множество зарядов, суммарная напряжённость определятся сложением напряжённостей, созданных каждым зарядом, по правилам сложения векторов.
2) Электрический потенциал. Чтобы переместить заряд против действующей на него электрической силы необходимо выполнить работу. Эта работа не зависит от пути перемещения заряда в электрическом поле, но зависит от начального и конечного положения заряда.
Если заряд перемещается из одной точки в другую против электрической силы, его потенциальная электростатическая энергия увеличивается. Электрический потенциал в любой точке равен электростатической потенциальной энергии Wp, которую имеет положительный заряд q в этой точке: φ = Wp/q (4).
Можно также сказать, что электрический потенциал в точке равен работе, которую необходимо совершить против электрических сил, чтобы переместить положительный заряд из данной точки на большое расстояние, где потенциал электрического поля равен нулю. Электрический потенциал является скалярной величиной и измеряется в вольтах (В).
Напряжённость электрического поля является отрицательным градиентом электрического потенциала - показателя изменения потенциала с расстоянием x: E→ = - dφ/dx. С помощью приборов можно измерить разность потенциалов, но не напряжённость поля. Последняя может быть вычислена, если использовать зависимость между E→ и Δφ : где Δφ = E·l - расстояние между двумя токами электрического поля.
Мембранный потенциал покояКаждая клетка превращает часть своей метаболической энергии в электростатическую энергию. Источником электрического поля клетки является плазматическая мембрана. Существует разность потенциалов между внутренней и внешней поверхностями плазматической мембраны. Эта разность потенциалов называется мембранным потенциалом.
Разность потенциалов между внутренней и внешней средами клетки может измеряться непосредственно и довольно точно. Для этого используют микроэлектрод, представляющий собой стеклянную микропипетку с диаметром кончика до 1мкм, заполненную концентрированным раствором KCl. Микроэлектрод подключают к усилителю напряжения регистрирующего устройства. Можно измерять мембранный потенциал мышечных, нервных клеток или клеток других тканей. Другой электрод (референтный) установлен на поверхности ткани.
Когда кончик микроэлектрода находится вне клетки, его потенциал по отношению к референтному электроду равен нулю. Если конец электрода погружают в клетку, прокалывая плазматическую мембрану, разность потенциалов резко становится отрицательной. На шкале измерительного устройства регистрируется разность потенциалов между внутренней и внешней средами клетки. Эта разность потенциалов называется трансмембранной, или мембранным потенциалом.
Если клетка находится в состоянии покоя, её мембранный потенциал имеет отрицательное значение и устойчивую величину. Обычно его называют мембранным потенциалом покоя. Мембранный потенциал покоя клеток различных тканей составляет от - 55 милливольт (мВ) до -100 мВ.
При определенных физиологических условиях могут происходить изменения мембранного потенциала. Изменения его в положительном направлении называется деполяризацией плазматической мембраны. Смещение мембранного потенциала в отрицательном направлении называется гиперполяризацией.
Биофизические основы мембранного потенциала покояЭлектрические явления в плазматической мембране определяются распределением ионов между внутренней и внешней сторонами мембраны. Из химического анализа известно, что концентрация ионов внутриклеточной жидкости сильно отличается от концентрации ионов во внеклеточной жидкости. Термин "внеклеточная жидкость" имеет отношение ко всем жидкостям вне клеток (межклеточное вещество, кровь, лимфа и т.п.). В таблице представлены концентрации основных ионов в мышечных клетках млекопитающих и внеклеточной жидкости (миллимоли на литр).
 |
Существуют значительные различия между концентрацией основных ионов внутри и вне клетки. Внеклеточная жидкость имеет высокую концентрацию ионов натрия и хлора. Внутриклеточная жидкость имеет высокую концентрацию калия и различных органических анионов (A-) (заряженные группы белков).
Различие между концентрациями натрия и калия во внеклеточной и внутриклеточной жидкостях обусловлены деятельностью натрий-калиевого насоса, который выкачивает за один цикл 3 иона натрия из клетки и закачивает 2 иона калия в клетку против электрохимического градиента указанных ионов. Основная функция натрия-калия насоса - поддержание различия концентраций ионов натрия и калия по обе стороны плазматической мембраны.
В состоянии покоя проницаемость плазматической мембраны для ионов калия значительно превышает проницаемость мембраны для ионов натрия. В нервных клетках соотношения проницаемости соответствующих ионов составляет 1:0,04.
Этот факт дает возможность объяснять существование мембранного потенциала покоя.
Ионы калия стремятся покинуть клетку из-за их высокой внутренней концентрации. При этом перемещения через мембрану внутриклеточных анионов из-за их больших размеров не происходит. Незначительное поступление ионов натрия внутрь клетки также не компенсирует выход ионов калия наружу, так как проницаемость мембраны в покое для ионов натрия мала.
Следовательно, снаружи клетка приобретает дополнительно положительный заряд и внутри остаётся избыток отрицательного заряда.
Диффузия калия через мембрану - процесс ограниченный. Ионы калия, проникающие через мембрану, создают электрическое поле, которое задерживает диффузию других ионов калия. По мере выхода из клетки калия электрическое поле нарастает и, в конечном итоге, напряжённость достигает такого значения, когда поток калия через мембрану прекращается. Состояние, при котором поток ионов по их концентрационному градиенту уравновешивается мембранным потенциалом, называется состоянием электрохимического равновесия ионов. Величина такого мембранного потенциала равновесия определяется уравнением Нернста (при этом считают, что мембрана проницаема только для одного вида ионов):
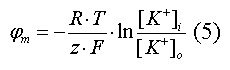 |
R - универсальная газовая постоянная, T - термодинамическая температура, z - электрический заряд иона, F - постоянная Фарадея, [K+]i и [K+]o - внутриклеточная и внеклеточная концентрации ионов калия соответственно.
Вычисления, основанные на уравнении Нернста, указывают, что внутренняя и внешняя концентрация иона хлора также соответствует состоянию электрохимического равновесия, но концентрация натрия далека от равновесия с мембранным потенциалом мембраны.
Уравнение Нернста показывает, что концентрационный градиент ионов калия определяет величину мембранного потенциала покоя только в первом приближении. Рассчитанные величины мембранного потенциала совпадают с экспериментально полученными только при высокой концентрации калия вне клетки.
Более точная величина мембранного потенциала покоя может быть вычислена из уравнения Гольдмана-Ходжкина, в котором учитываются концентрации и проницаемость мембраны для трёх основных ионов внутри- и внеклеточной жидкостей:
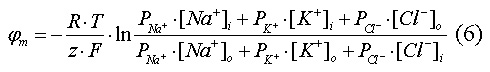 |
Также в поддержании мембранного потенциала покоя участвует непосредственно натрий-калий насос, выкачивая три иона натрия из клетки и закачивая лишь два иона калия. В результате мембранный потенциал покоя становится более отрицательным, чем был бы, если бы создавался только пассивным перемещением ионов через мембрану.
Потенциал действияЕсли через мембрану нервной или мышечной клетки проходит кратковременный электрический ток, то мембранный потенциал подвергается последовательным изменениям, которые специфичны и уникальны для возбудимых клеток. Возбудимые ткани можно стимулировать также механическими или химическими средствами, но в экспериментальной работе, как правило, используются электрические стимулы.
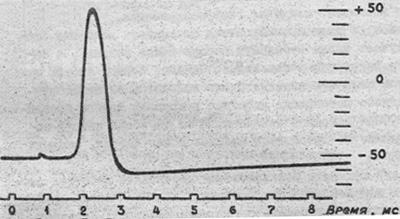 Рис. 1. Потенциал действия нервной клетки. Рис. 1. Потенциал действия нервной клетки.
Потенциал действия - быстрое колебание величины мембранного потенциала, вызванное действием на возбудимую клетку электрического или другого раздражителей.
На рис. 1 показан потенциал действия нервной клетки, записанный с помощью микроэлектрода. Если к клетке прикладывают кратковременный электрический стимул, мембранный потенциал уменьшается быстро до нуля. Это отклонение характеризуют как фазу деполяризации. В течение короткого времени внутренняя среда клетки становится электроположительна по отношению к наружней (фаза реверсии мембранного потенциала, или овершут). Затем мембранный потенциал возвращается к уровню мембранного потенциала покоя (этап реполяризации) (рис.2.).
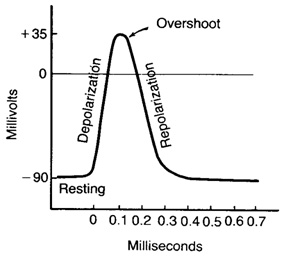 Рис. 2. Фазы потенциала действия Рис. 2. Фазы потенциала действия
Длительность потенциала действия составляет от 0,5 до 1 миллисекунды в больших нервных клетках и несколько миллисекунд в клетках скелетных мышц. Общая амплитуда - почти 100 - 120мВ, отклонение от нулевой линии - около 30-50мВ.
Потенциал действия играет ведущую роль в обработке информации в нервной системе. Он имеет постоянную амплитуду, которая не является вероятностной величиной. Это имеет большое значение в обработке информации нервной системой. Кодирование интенсивности раздражения осуществляется числом потенциалов действия и частотой, с которой потенциалы действия следуют друг за другом.
Биофизические основы потенциала действияПотенциал действия возникает из-за специфических изменений ионной проницаемости в плазматической мембране. Английский физиолог Ходжкин показал, что основной механизм потенциала действия состоит в кратковременном и очень специфическом изменении проницаемости мембраны для ионов натрия. Ионы натрия при этом поступают в клетку до момента, пока мембранный потенциал не достигнет потенциала электрохимического равновесия ионов натрия.
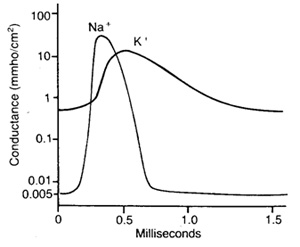 Рис. 3. Изменение проницаемости мембраны для ионов натрия и калия во время потенциала действия Рис. 3. Изменение проницаемости мембраны для ионов натрия и калия во время потенциала действия
Проницаемость мембраны для натрия при действии на клетку электрического стимула возрастает приблизительно в 500 раз и становится значительно больше, чем проницаемость мембраны для ионов калия. В клетке резко повышается концентрация ионов натрия. В результате мембранный потенциал принимает положительное значение, и поток ионов натрия в клетку замедляется.
Во время возникновения потенциала действия происходит деполяризация плазматической мембраны. Быстрая деполяризация мембраны под действием электрического стимула вызывает увеличение её проницаемости для ионов натрия. Возросшее поступление ионов натрия в клетку усиливает деполяризацию мембраны, что, в свою очередь, вызывает дальнейшее увеличение проницаемости мембраны для натрия и т.д.
Но величина мембранного потенциала при деполяризации не достигает уровня потенциала электрохимического равновесия ионов натрия. Причиной этому является снижение проницаемости мембраны для ионов натрия из-за инактивации натриевого трансмембранного переноса. Этот процесс резко уменьшает проницаемость мембраны для ионов натрия и останавливает наплыв натрия в клетку.
В этот момент происходит увеличение проницаемости мембраны для ионов калия, что приводит к быстрому снижению величины мембранного потенциала к уровню потенциала покоя. Проницаемость мембраны для ионов калия также снижается до своего нормального значения. Таким образом, инактивация входящего натриевого тока и повышение проницаемости мембраны для ионов калия (выходящий ток) ограничивают длительность потенциала действия и приводят к реполяризации мембраны.
Таким образом, в течение потенциала действия некоторое количество ионов натрия поступают в клетку. Но это количество достаточно небольшое. Изменение концентрации ионов в больших нервных клетках составляет лишь около 1/300000 начальной величины.
Основной механизм изменений проницаемости мембраны обусловлен событиями в натриевых и калиевых каналах мембраны. Состояние их ворот управляется величиной мембранного потенциала. Натриевые каналы имеют два типа ворот. Один из них, называемые активационными воротами закрыты в состоянии покоя и открываются при деполяризации мембраны. Поступление ионов натрия в клетку вызывает открытие всё большего числа активационных ворот. Второй тип ворот натриевых каналов - инактивационные при усиливающейся деполяризации мембраны постепенно закрываются, что останавливает приток натрия в клетку. Деполяризация мембраны также служит причиной открытия дополнительного числа калиевых каналов, в результате чего увеличивается проницаемость мембраны для ионов калия и происходит реполяризация мембраны.
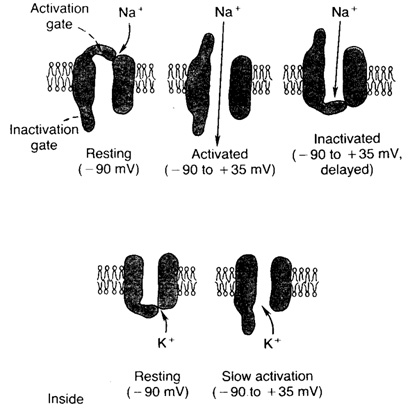 Рис. 4. Изменение состояния натриевых и калиевых каналов мембраны в зависимости от величины мембранного потенциала Рис. 4. Изменение состояния натриевых и калиевых каналов мембраны в зависимости от величины мембранного потенциала
Распространение потенциала действияПотенциал действия распространяется вдоль мембраны нервной и мышечной клеток без уменьшения амплитуды с расстоянием. Этот процесс обусловлен кабельными свойствами плазматической мембраны, т.е. способностью проводить электрический ток на небольшие расстояния. Локальный электрический ток течет в клетку в активной области (где возникает потенциал действия) и из клетки - в смежной неактивной зоне. Эти ионные токи вызывают некоторые изменения мембранного потенциала в зоне, прилегающей к месту возникновения потенциала действия.
Циклический локальный ток снижает заряд мембраны в неактивной зоне и деполяризует её. Если деполяризация достигает порогового уровня, то возрастает проницаемость мембраны для ионов натрия и возникает потенциал действия. Таким образом потенциал действия распространяется вдоль нервных и мышечных волокон с постоянной скоростью.
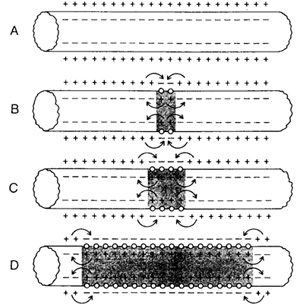 Рис. 5. Распространение потенциала действия вдоль мембраны нервного волокна Рис. 5. Распространение потенциала действия вдоль мембраны нервного волокна
Скорость распространения потенциала действия в нервных волокнах зависит от их диаметра. Она максимальна в наиболее толстых волокнах, достигая около 100 метров в секунду.
СМОТРИТЕ ТАКЖЕ:
Социальные комментарии Cackle
|