|
|
Физика - одна из самых удивительных наук! Физика столь интенсивно развивается, что даже лучшие педагоги сталкиваются с большими трудностями, когда им надо рассказать о современной науке. Данный ресурс поможет эффективно и интересно изучать физику. Учите физику! |
|||
| полезное |
|---|
| Смешные анекдоты о физике
Готовые шпоры по физике Физика в жизни Ученые и деньги Нобелевские лауреаты Фото Видео Карта сайта |
| На заметку |
|---|
| Если вам понравился сайт, предлагаем разместить нашу кнопку |
| Дополнительно |
|---|
|
Компьютерные программы по физике 
Физика и юмор 
Онлайн тестирование по физике 
|
Нервные механизмы зрения
Одной из основных тем этой главы является взаимосвязь и взаимоинформация отдельных частей глаза. Давайте рассмотрим сложный глаз краба-мечехвоста, над которым было проделано довольно много опытов. Прежде всего нужно понять, какого сорта информация может передаваться по нервам. По нерву передается нечто вроде возмущения электрической природы, которое может быть легко зарегистрировано. Это некое волнообразное возмущение, которое бежит по нерву и вызывает на другом его конце какой-то эффект. Информацию переносит длинный отросток нервной клетки, называемый аксоном, и если один конец аксона возбужден, то по нему бежит «импульс». Далее, если по нерву уже проходит один импульс, то за ним не может немедленно последовать второй. Все импульсы имеют одну и ту же величину, так что, когда нерв сильно возбужден, это вовсе не означает, что по нему бежит больший импульс, а просто увеличивается число импульсов в 1 сек. Величина же импульса определяется нервным волокном. Это важно усвоить, чтобы понять, что произойдет дальше.
Однако тот факт, что существует такое «усиление» контура, известен уже давно. Это действительно замечательная вещь, которая неоднократно обсуждалась психологами. Чтобы изобразить предмет, нам достаточно нарисовать лишь его контур. Ведь мы привыкли видеть картины, на которых изображены одни только контуры! Но что же такое контур? Ведь это просто граница между темным и светлым или между одним цветом и другим. В самом деле, это вовсе не что-то определенное. Можно думать, что угодно, но никакой линии вокруг предмета нет! Нет, все это только наша выдумка. Теперь мы начинаем понимать, почему нам достаточно контура, чтобы представить себе весь предмет. По-видимому, наши глаза тоже работают, подобно глазам мечехвоста, разумеется, гораздо более сложно, но все же аналогично. Наконец, я вкратце опишу более сложные опыты, очень красивые, и трудные, которые были проделаны над лягушкой. При выполнении их в зрительный нерв лягушки вводились миниатюрные искусно сделанные нитеобразные зонды и измерялись сигналы, идущие вдоль одного определенного аксона; точно так же как в случае с крабом-мечехвостом, было обнаружено, что информация зависит не просто от одной точки глаза, а является суммой информации, полученных от нескольких частей. Наиболее современная картина операций на глазе лягушки выглядит следующим образом. Можно найти четыре типа различных зрительных нервных волокон, в том смысле, что существуют четыре различных сорта ответных реакций. В этих экспериментах не было вспышек света: лягушка не замечает таких вещей. Она просто сидит, и глаза ее неподвижны до тех пор, пока листок лилии не начнет двигаться. Тогда глаза лягушки движутся как раз так, чтобы изображение оставалось в поле зрения. Однако сама по себе лягушка глазами не ворочает и не ищет, куда девался объект. Если в поле ее зрения движется что-то, напоминающее небольшое насекомое (нужно, чтобы она могла видеть нечто маленькое, движущееся на неподвижном фоне), то обнаруживаются четыре различных сорта нервных волокон, отвечающих на это раздражение. Их свойства приведены в табл. 36.1. Длительное (нестираемое) обнаружение края
означает, что если мы внесем предмет с резким краем в поле зрения лягушки, то в тех фоторецепторах, мимо которых он движется, возникает множество импульсов, которые переходят затем в редкие импульсы, продолжающиеся до тех пор, пока край находится в поле зрения, даже если он стоит на месте. После выключения света импульсы прекращаются. Если снова включить свет и край предмета по-прежнему находится в поле зрения, то импульсы возникают снова. Они не исчезают. Другой сорт волокон очень похож на первый, но с тем исключением, что они не работают, если край прямой. Нужно, чтобы край был изогнутым! Насколько сложной должна быть система взаимных связей сетчатки глаза лягушки, чтобы видеть движение изогнутой поверхности! Более того, если эти волокна чем-то возбуждены, то возбуждение не может держаться так же долго,как в первом случае, и если мы выключим свет и включим его снова, то импульсы не возобновятся. Впрочем, это зависит от движения выпуклой поверхности. Глаз видит ее движение и помнит, где она находится, но если мы на момент выключим свет, то глаз просто забывает о ней и больше ее не видит. Следующим типом является регистрация изменения контраста. Если край надвигается или отодвигается, то сигналы есть. Но если предмет стоит, то никаких сигналов вообще нет. Затем есть «регистратор тусклости». Если интенсивность света уменьшается, то возникают импульсы, если она неизменна — импульсы прекращаются: регистратор работает, только когда свет тускнеет. И наконец, есть несколько волокон, которые служат регистраторами темноты. Самое удивительное, что они беспрестанно «стреляют»! Если свет усиливается, «выстрелы» становятся более редкими, а если ослабевает, то, наоборот, «огонь» становится более частым, но он не прекращается ни на секунду. В темноте же они «строчат», как сумасшедшие, напоминая постоянно: «Тьма! Тьма! Тьма!» Все эти реакции кажутся слишком сложными, чтобы их можно было как-то классифицировать. Подозрительно даже, правильно ли истолкованы эксперименты. Но самое интересное, что эти же классы очень отчетливо выделяются самой анатомией лягушки! После того как волокна были расклассифицированы (очень важно, что это было сделано после), другие измерения обнаружили, что скорости импульсов, бегущих по различным волокнам, не одинаковы. Так что был найден другой независимый способ определения сорта волокна! Еще один интересный вопрос: насколько велика анализирующая область, связанная с каким-то одним волокном? Ответ оказался различным для разных классов волокон.
Все, что мы узнали, можно кратко сформулировать так: по-видимому, имеются три сорта пигментов. Может быть множество различных сортов рецепторов, в которые эти пигменты входят в различных пропорциях, однако множество внутренних связей позволяет складывать и вычитать эффекты отдельных нервных клеток Таким образом, прежде чем мы действительно поймем цветовое зрение, необходимо понять конечный этап ощущение зрения вообще. Это все еще открытый вопрос, но исследования с микроэлектродами, возможно, дадут нам в конце концов дополнительные сведения о том, как же мы видим цвета. СМОТРИТЕ ТАКЖЕ:Социальные комментарии Cackle |
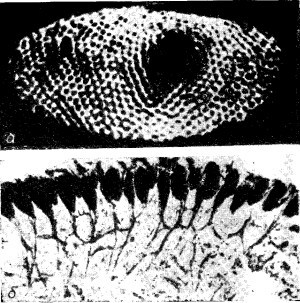 На фиг. 36.11,а показан сложный глаз краба-мечехвоста; в нем всего лишь около тысячи омматидиев. Фиг. 36.11, б представляет собой поперечный разрез этой системы. Видны отдельные омматидии и нервные волокна, соединяющие их с мозгом. Но обратите внимание, что даже у этого краба имеются внутренние связи. Они, конечно, гораздо менее сложные, чем в глазе человека, но именно это-то и дает нам возможность изучить подобные связи на простом примере.
На фиг. 36.11,а показан сложный глаз краба-мечехвоста; в нем всего лишь около тысячи омматидиев. Фиг. 36.11, б представляет собой поперечный разрез этой системы. Видны отдельные омматидии и нервные волокна, соединяющие их с мозгом. Но обратите внимание, что даже у этого краба имеются внутренние связи. Они, конечно, гораздо менее сложные, чем в глазе человека, но именно это-то и дает нам возможность изучить подобные связи на простом примере.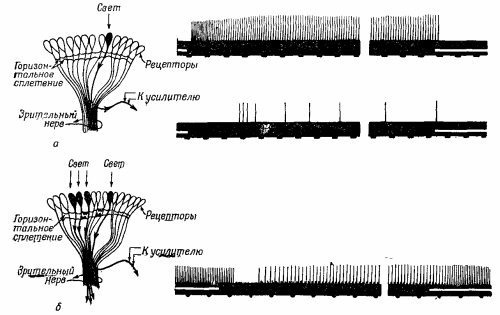 Давайте рассмотрим такой опыт: наложим на зрительный нерв нашего краба небольшие электроды и осветим только один омматидий; это легко можно сделать с помощью линз. Если мы в какой-то момент времени t0 включим свет и будем измерять возникающие электрические импульсы, то увидим, что после небольшой задержки последует быстрая серия разрядов, частота которых постепенно будет уменьшаться, пока не достигнет какой-то равномерности (фиг. 36.12,а). После выключения света разряды прекращаются. Интересно, что если усилитель остается связанным с тем же нервом, а мы направим свет на другой омматидий, то ничего не произойдет, сигналов не будет.
Давайте рассмотрим такой опыт: наложим на зрительный нерв нашего краба небольшие электроды и осветим только один омматидий; это легко можно сделать с помощью линз. Если мы в какой-то момент времени t0 включим свет и будем измерять возникающие электрические импульсы, то увидим, что после небольшой задержки последует быстрая серия разрядов, частота которых постепенно будет уменьшаться, пока не достигнет какой-то равномерности (фиг. 36.12,а). После выключения света разряды прекращаются. Интересно, что если усилитель остается связанным с тем же нервом, а мы направим свет на другой омматидий, то ничего не произойдет, сигналов не будет.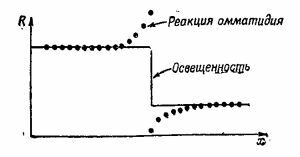 Проделаем теперь другой опыт: осветим первый омматидий и получим те же импульсы, но если теперь мы направим свет еще на другой соседний омматидий, то на короткое время импульсы прекращаются, после чего импульсы «побегут» снова, но с гораздо меньшей частотой (фиг. 36.12,6). Оказывается, что импульсы, возникающие во втором омматидий, затормаживают импульсы первого! Другими словами, хотя каждый нерв и несет информацию о своем омматидий, количество этой информации подавляется сигналами от другого омматидия. Например, когда более или менее равномерно освещен весь глаз, то сигнал, пришедший от любого отдельного омматидия, будет относительно слабым, ибо он подавлен множеством других сигналов. Торможение аддитивно, т. е. если мы осветим несколько соседних омматидиев, торможение будет очень сильным. Торможение оказывается большим, если омматидии расположены ближе, но если они достаточно удалены друг от друга, то торможение практически сводится к нулю. Таким образом, торможение аддитивно и зависит от расстояния. Это первый пример, когда информация от различных частей глаза перерабатывается в нем самом. Если подумать немного, то можно понять, что этот механизм предназначен для усиления контраста на краях объекта, ибо если часть предмета освещена, а часть нет, то омматидии, направленные на освещенную область, дают импульсы, которые затормаживаются всеми соседними омматидиями, видящими освещенную область, так что они относительно слабы. С другой стороны, омматидии, видящие границу световой области и дающие «белый» сигнал, хотя и подавлены своими соседями, но тех не столь уж много, ибо некоторые из них совсем темные ( не видят света), в результате чего сигнал оказывается более сильным. В итоге получается кривая, подобная изображенной на фиг. 36.13. Краб как бы видит «усиленный» контур.
Проделаем теперь другой опыт: осветим первый омматидий и получим те же импульсы, но если теперь мы направим свет еще на другой соседний омматидий, то на короткое время импульсы прекращаются, после чего импульсы «побегут» снова, но с гораздо меньшей частотой (фиг. 36.12,6). Оказывается, что импульсы, возникающие во втором омматидий, затормаживают импульсы первого! Другими словами, хотя каждый нерв и несет информацию о своем омматидий, количество этой информации подавляется сигналами от другого омматидия. Например, когда более или менее равномерно освещен весь глаз, то сигнал, пришедший от любого отдельного омматидия, будет относительно слабым, ибо он подавлен множеством других сигналов. Торможение аддитивно, т. е. если мы осветим несколько соседних омматидиев, торможение будет очень сильным. Торможение оказывается большим, если омматидии расположены ближе, но если они достаточно удалены друг от друга, то торможение практически сводится к нулю. Таким образом, торможение аддитивно и зависит от расстояния. Это первый пример, когда информация от различных частей глаза перерабатывается в нем самом. Если подумать немного, то можно понять, что этот механизм предназначен для усиления контраста на краях объекта, ибо если часть предмета освещена, а часть нет, то омматидии, направленные на освещенную область, дают импульсы, которые затормаживаются всеми соседними омматидиями, видящими освещенную область, так что они относительно слабы. С другой стороны, омматидии, видящие границу световой области и дающие «белый» сигнал, хотя и подавлены своими соседями, но тех не столь уж много, ибо некоторые из них совсем темные ( не видят света), в результате чего сигнал оказывается более сильным. В итоге получается кривая, подобная изображенной на фиг. 36.13. Краб как бы видит «усиленный» контур.
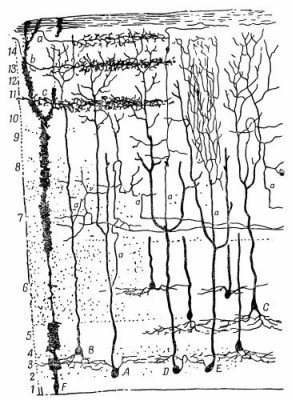 На фиг. 36.14 показана поверхность так называемой покрышки мозга лягушки. Сюда приходят волокна из зрительного нерва. Все эти нервные волокна связаны с различными слоями покрышки. Слоистое строение ее напоминает строение сетчатки (это один из фактов, говорящих нам о том, что сетчатка и мозг весьма похожи друг на друга). Если теперь взять электрод и постепенно перемещать его вниз по слоям, то можно определить, где оканчиваются разные типы зрительных волокон. Опыт дает очень красивый и удивительный результат: оказывается, что различные сорта волокон оканчиваются в различных слоях! В первом слое оканчивается первый тип волокон, во втором — второй; третий и пятый оканчиваются в одном и том же слое, а глубже всех проникает четвертый тип. (Вас не должно удивлять, что номера их почти совпали с номерами слоев! Именно поэтому они и пронумерованы таким образом, в ранних работах они нумеровались иначе!)
На фиг. 36.14 показана поверхность так называемой покрышки мозга лягушки. Сюда приходят волокна из зрительного нерва. Все эти нервные волокна связаны с различными слоями покрышки. Слоистое строение ее напоминает строение сетчатки (это один из фактов, говорящих нам о том, что сетчатка и мозг весьма похожи друг на друга). Если теперь взять электрод и постепенно перемещать его вниз по слоям, то можно определить, где оканчиваются разные типы зрительных волокон. Опыт дает очень красивый и удивительный результат: оказывается, что различные сорта волокон оканчиваются в различных слоях! В первом слое оканчивается первый тип волокон, во втором — второй; третий и пятый оканчиваются в одном и том же слое, а глубже всех проникает четвертый тип. (Вас не должно удивлять, что номера их почти совпали с номерами слоев! Именно поэтому они и пронумерованы таким образом, в ранних работах они нумеровались иначе!)При использовании материалов сайта ссылка на www.all-fizika.com обязательна.